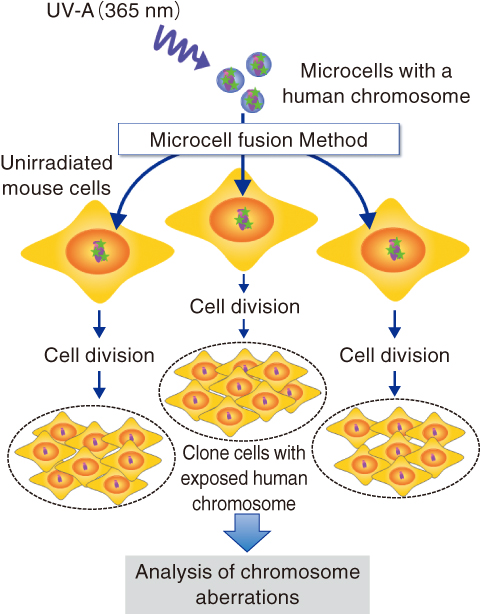
Fig.3-10 Transfer of damaged DNA by a microcell fusion method
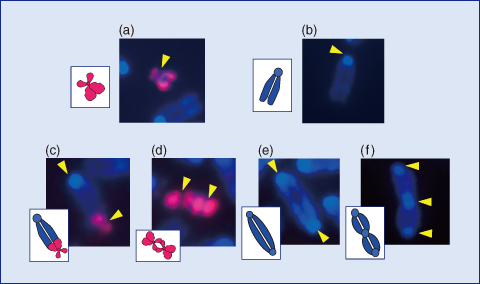
Fig.3-11 Typical chromosome aberrations microscopically observed
As one of the genetic effects through multiple repetitions of cell division, chromosome aberrations are occasionally induced in the progeny cells after ionizing irradiation. This is known to be “genetic instability,”which is strongly involved in the induction of late radiation effects, such as carcinogenesis. The detailed mechanism behind it, however, has not yet been fully understood.
In this study, we focused on DNA damage as a trigger of the instability, particularly DNA base damage, which might persist even after repeated cell divisions. The bases normally work as elementary units of genetic code in DNA, and their alteration is thought to raise mutation frequencies. In addition to DNA damage, irradiation could also induce damage to cytoplasmic organelles. To exclude the cytoplasmic effects, we applied a microcell fusion method to observe genetic instabilities induced only by DNA base damage.
Sub-cellular small structures called microcells, including one human chromosome, were irradiated and then transferred into unirradiated normal mouse cells (Fig.3-10). The human chromosome is considerably different from those of the mouse cell, such that the exposed human chromosome was easily distinguished from unirradiated mouse chromosomes by means of microscopic observations. It was confirmed beforehand that normal chromosome transfer did not induce any genetic instabilities. Ultraviolet light (wavelength: 365 nm), known as UV-A, was used as the source of the irradiation, because it has been known to preferentially induce the same sort of DNA base lesions that are induced by ionizing radiation. After the chromosome transfer, the cells were cultured for 20 days or a month to obtain various clonal cell lines that stably undergo cell division.
The analysis of the chromosomes in the clones revealed that certain types of aberrations, such as fusions between normal mouse chromosomes, were frequently induced, in addition to fusions between the human chromosomes, or between human and mouse chromosomes (Fig.3-11). Furthermore, it was observed that the number of chromosomes in the clone cells abnormally increased from the normal number of 43 to twice that number or more.
These evidences indicate that a novel mechanism of signal transfer from damaged to normal chromosomes could result in the genetic instability in the unirradiated cells. Detailed investigation is expected to reveal the induction process of radiation carcinogenesis in the low dose region in the near future.
The present study was partly supported by the Ministry of Education, Culture, Sports, Science and Technology of Japan (MEXT) KAKENHI Grant-in-Aid for Young Scientists (B) (No.20710045).

