6-4 |
DNA修復遺伝子のスイッチを入れるユニークな蛋白質を発見 |
 |
||
|
 |
||
|
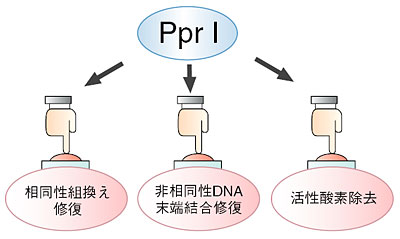
| 放射線抵抗性細菌デイノコッカス・ラジオデュランスは、放射線照射によって生じるDNAの損傷を克服する能力が特に優れています。DNA損傷は、放射線照射や紫外線照射といった外的要因の他に、呼吸や代謝の過程で生じる活性酸素やDNAの複製ミス等の内的要因によっても生じます。ラジオデュランスでDNA損傷が生じた後、特定の遺伝子群の発現の活性化現象があることが分かっていましたが、どのようにして遺伝子発現の活性化が起こるのかが謎でした。 通常、ラジオデュランスのDNA修復能欠損株に正常株のDNAを導入し、DNA修復能を正常に復帰させることのできるDNA領域を限定することで、放射線感受性の原因遺伝子を特定することができます。しかしながら、DNA修復能欠損株KH8401は、DNAを導入しなくても一部の細胞が高頻度で放射線耐性になるという性質(図6-8)があるので、従来の方法を用いて放射線感受性の原因遺伝子を特定することが不可能でした。 私たちは、KH8401株のユニークな性質が、ゲノムDNAの中を自由に動き回ることのできるトランスポゾンによって引き起こされているのではないかと考え、KH8401株と正常株のゲノムにおけるトランスポゾンの分布を比較しました。その結果、KH8401株のみにトランスポゾンの転移挿入が認められるDNA領域を見出し、挿入部位の同定により放射線感受性の原因遺伝子を同定しました。この遺伝子から作られる蛋白質の性質を調べた結果、相同性組換え修復や非相同性DNA末端結合修復に係わる遺伝子と、活性酸素除去に係わるDNA防御遺伝子の発現を活性化する主要スイッチ蛋白質であることが分かり、「DNA修復を促進する多面的蛋白質群の誘導(inducer of pleiotropic proteins promoting DNA repair)」に係わることから、PprIと命名しました(図6-9)。 PprI蛋白質によって制御されている遺伝子群を調べることで、ラジオデュランスの放射線応答機構の全容を解明することができると考えられます。また、PprI蛋白質を有効利用することによって、広範な線量域で使用可能な生物線量計を開発することが可能となります。 |
| ●参考文献 Y. Hua et al., PprI: A General Switch Responsible for Extreme Radioresistance of Deinococcus radiodurans, Biochem. Biophys. Res. Commun., 306(2), 354 (2003). |
| ご覧になりたいトピックは左側の目次よりお選びください。 |  |
| たゆまざる探究の軌跡−研究活動と成果2004
Copyright(c) 日本原子力研究所 |