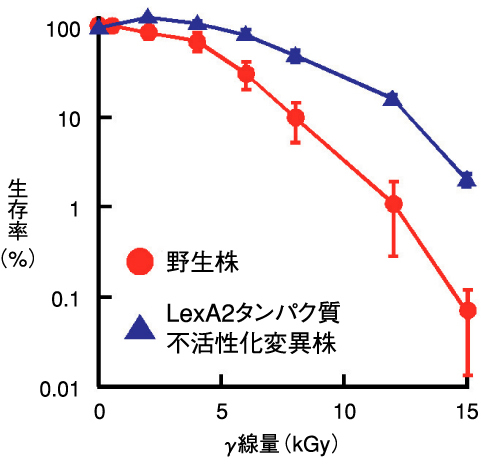
図4-10 γ線に対する生存率
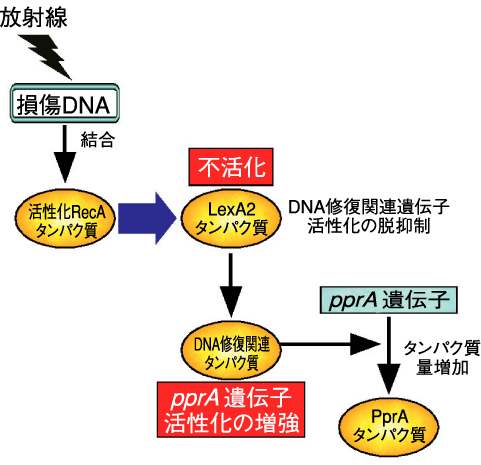
図4-11 ラジオデュランスの放射線応答機構
微生物の放射線応答機構は、1970年代から大腸菌で研究が盛んに行われており、放射線照射後にDNA修復タンパク質群の細胞内生産の増加を促す仕組みがあることが分かっています。RecAタンパク質(Recombinase「組換え酵素」に由来)とLexAタンパク質(Locus for X-raysensitivity「X線感受性に関する遺伝子領域」に由来)がこの応答の中心的役割を担っています。一方、私たちは、放射線抵抗性細菌デイノコッカス・ラジオデュランス(ラジオデュランス)の放射線応答機構にRecAタンパク質が関与するが、LexAタンパク質は関与していないことを明らかにしました。また、私たちは、DNA修復促進タンパク質PprA(Pleiotropic protein promoting DNA repair「DNA修復を促進する多面的タンパク質」に由来)を発見し、それが放射線耐性機構に重要であることを明らかにしてきましたが、放射線応答機構におけるPprAタンパク質とRecAタンパク質の関係は不明でした。
ラジオデュランスはLexAタンパク質の他に、アミノ酸配列が類似したタンパク質(LexA2と命名)を持っています。そこで、私たちはバイオインフォマティクスと分子生物学の連携によって、LexA2タンパク質の不活性化によりラジオデュランスの放射線耐性が向上することを明らかにしました(図4-10)。ラジオデュランスの全ゲノムDNA塩基配列は、1999年に米国ゲノム研究所が解読済みです。しかし、解読した配列のlexA2 遺伝子相当領域には間違いがあり、これまでこの間違った配列をもとに研究が行われてきました。このため、私たちはlexA2 遺伝子の正しいDNA塩基配列を決定し、これに基づき正しいアミノ酸配列を持つLexA2タンパク質の性質を詳細に調べ、その結果、次のことを明らかにしました。RecAタンパク質は損傷DNAと結合することで活性化します。活性化RecAタンパク質とLexA2タンパク質が相互作用することでLexA2タンパク質が不活性化されます。これにより、DNA修復関連遺伝子活性化の脱抑制が起こり、pprA 遺伝子の活性を増強することで、最終的にPprAタンパク質の細胞内生産が増加するのです(図4-11)。この一連の出来事が、LexA2タンパク質を不活性化した変異株の放射線耐性が向上する一因であると考えられました。
現在、PprAタンパク質は高効率のDNA修復試薬として製品化されていますが、今回の研究成果に基づき、試薬製造過程におけるPprAタンパク質生産効率の向上への応用が可能となります。更に、DNA修復タンパク複合体の立体構造解析を行い、タンパク質相互作用の詳細な分子機構を明らかにすることで、医療の高度化に役立つ新たなDNA修復タンパク複合体による創薬の開発が期待されます。

