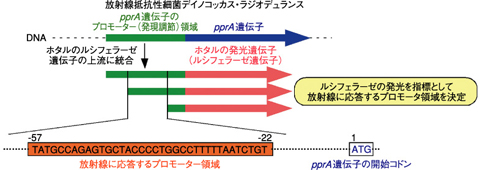
図4-14 pprA遺伝子の放射線応答プロモーター領域の同定
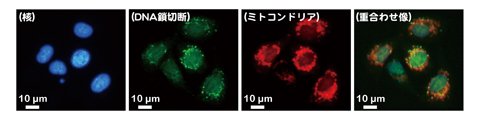
図4-15 γ線照射後の哺乳動物細胞ミトコンドリア内DNA鎖切断
放射線抵抗性細菌デイノコッカス・ラジオデュランスは、放射線に対して著しく高い耐性を示します。これは、この菌が、放射線で誘発されるDNA損傷に対して優れた修復能力を持つことに起因しています。私たちは、初めてこの菌からpprA遺伝子を発見し、pprA遺伝子から作られるPprAタンパク質がDNAの鎖切断部位に特異的に結合し、DNA修復を促進する重要な機能をもつことを明らかにしてきました。しかし、放射線に応答したpprA遺伝子の活性化に必要なDNA領域(プロモーター領域)の同定が未解決である一方、PprAタンパク質の機能から期待されるDNA損傷の可視化検出技術への応用も確立されていませんでした。
そこで、プロモーター領域を同定するために、pprA遺伝子のプロモーターを含むDNA領域をホタルの発光遺伝子(ルシフェラーゼ遺伝子)の上流に結合させ、放射線照射によるルシフェラーゼの発光の変化を指標として、プロモーターによる遺伝子発現活性を測定しました。その結果、pprA遺伝子の上流−57から−22の領域が遺伝子の活性化に必要なプロモーター領域であることを明らかにしました(図4-14)。この放射線応答プロモーターの同定は、放射線抵抗性細菌のDNA修復ネットワーク機構の全容を解明するために、非常に有益な知見です。
一方、放射線に高い感受性を示す哺乳動物は、細胞内に生じたわずかなDNA鎖切断が細胞死にかかわります。したがって、哺乳動物の放射線影響を詳細に評価するためには、細胞内のDNA損傷の生成頻度や分布を直接的に測定することが重要です。これまでにも、哺乳動物細胞自身のタンパク質を利用して、細胞内に生じたDNA鎖切断を可視化検出する方法がありましたが、以前の検出方法では、DNAの初期損傷を検出するのが困難であるといった欠点がありました。そこで、この欠点を克服するために、放射線抵抗性細菌のPprAタンパク質が、DNA鎖切断部位に特異的に結合することを利用して、DNAの初期損傷を検出する方法を開発しました。この方法では、哺乳動物細胞のDNA鎖切断部位にPprAタンパク質を結合させ、更に蛍光標識したPprAタンパク質抗体を反応させることでDNAの初期損傷を可視化しています(図4-15)。この方法を改良し、検出感度を向上させることで、哺乳動物細胞の放射線影響の評価だけではなく、DNAに損傷を与える化学物質の遺伝毒性試験をはじめとする環境や医学分野での応用が可能と考えられます。