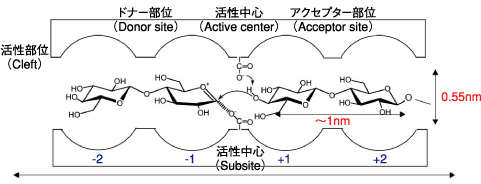
図6-8 酵素分子内の活性部位(クレフト)の模式図

図6-9 酵素重合により生成したポリマーの自己組織化構造の走査電子顕微鏡像(a)と火口からの火山灰の噴煙構造(b)との比較 、(c)は(b)の拡大像
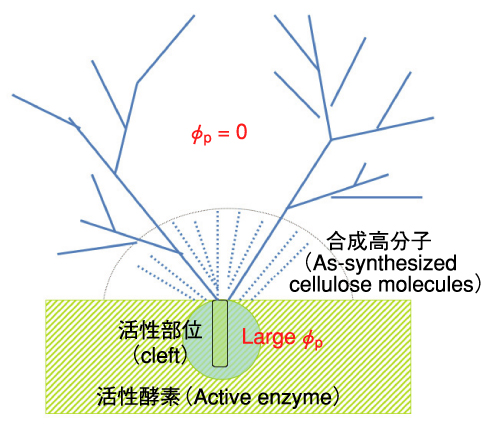
図6-10 酵素会合体の一部(緑色斜線)と反応溶液との界面
天然高分子の一つであるセルロースは、植物又はバクテリアの「特異な重合場」の中で生合成され、合成された高分子は“その場”でセルロース繊維へと自己組織化されます。この特異な重合場とは、植物,バクテリアが内蔵する酵素分子の活性部位(クレフト,cleft)であり、nmスケールの空間的に極めて「狭い反応場」であります(図6-8)。他方、有機合成化学法に基づいたセルロースの人工合成は1941年以来数多く試みられましたが、いずれも成功には至りませんでした。小林らは1991年、自ら合成した特殊なモノマー(図6-8左側中央)とセルラーゼという酵素分子を触媒として用いることによりセルロースの人工合成に初めて成功しました(J.Am.Chem.Soc.,vol.113,1991,p.3079-3084)。
私たちは、天然セルロースの生合成と自己組織化ついての生体の営みの謎に迫るために、より単純な小林らの人工合成系とJRR-3の中性子超小角散乱装置及び小角散乱装置を用い、合成過程・自己組織化過程のその場観察に世界で初めて挑戦しました。その結果、(1)酵素分子は反応溶液中で200nm以上の大きさの会合体を形成すること、(2)会合体の活性部位は、活性酵素(activeenzyme)1g当り約14kg、1個当り毎秒約5個のセルロース分子を生み出すこと、(3)セルロース分子は酵素会合体の周りに会合し、30nmから30μmの空間スケールにわたる表面フラクタル構造を形成することを明らかにできました(図6-9(a))。3桁にわたる自己相似パターンの実証は、フラクタル幾何学の分野で極めてまれで貴重です。
この造形の謎について以下の解釈を提案しました。クレフトの中でポリマーが重合成長すると、その周辺のポリマーの局所濃度φpは極めて高くなります。一方、反応溶液のポリマーの溶解度は極めて小さいので、溶液中でポリマーは、すぐに会合し結晶化します。そのため溶液とクレフト周辺のφpの差が極めて大きくなります(図6-10)。その結果合成されたポリマーはクレフト内から反応溶液中へと噴出し(図6-10)、ポリマー(図6-8右側中央)の成長末端はクレフト内の活性中心から引離されることになります。引離されたポリマーは重合停止し、活性部位は新たな重合反応を繰り返します。重合を停止したポリマーは反応溶液中で会合し、樹枝状多結晶体へと自己組織化します(図6-10)。このように、重合,拡散,重合停止,自己組織化という一連の過程を経て、図6-9(a)に象徴されるフラクタル構造が造形されるわけです。この自己組織化パターンは、空間スケールの1億倍の相違を除けば、火口からの噴煙の構造(図6-9(b),(c))と酷似しています。図6-9の二つの構造の類似性は酵素反応が秘める巨大な化学反応エネルギーを直感させてくれます。この成果は、生体系の造形の謎解きに関して、中性子ビーム利用研究が未来を拓く力を持つことを強く示唆するものです。

